phase plane of the wilson-cowan model
55 views
Skip to first unread message

Wer White
Jan 2, 2022, 8:29:57 AM1/2/22
to TVB Users
Dear TVBer,
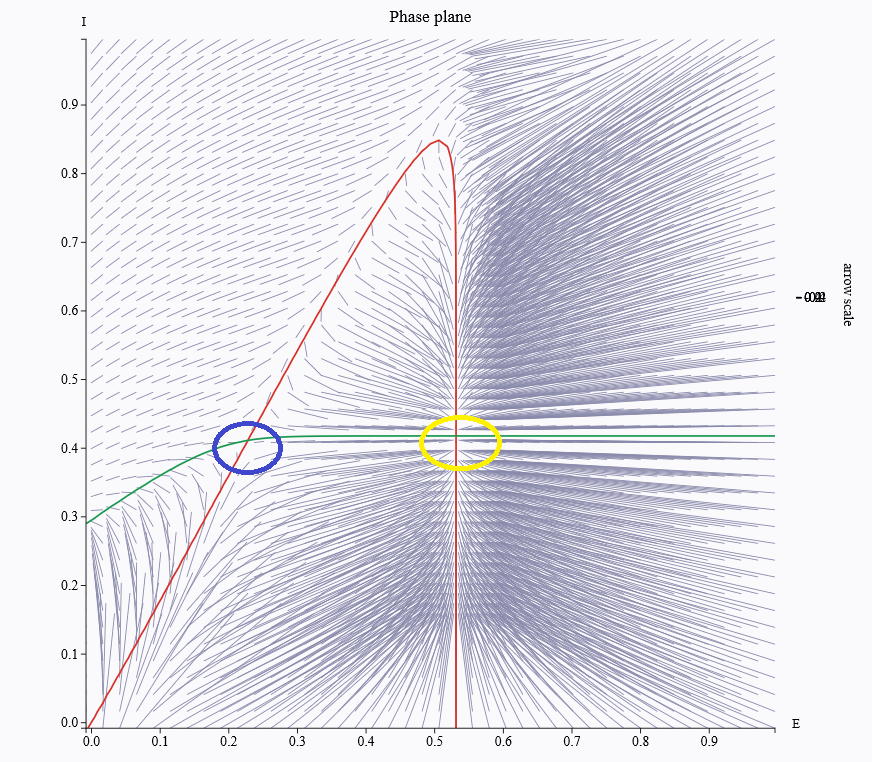
I'm still new to TVB and I'm puzzled when I tune parameters by phase plane analysis. I have some questions:
1. Is blue circle is a unstable point (bifurcation) and yellow circle is a stable point?
2. can i use these parameters corresponding to the figure?
3. i know fc appears at the
bifurcation point, does it mean that it appears at the blue circle?
Looking forward for your reply! Please forgive my stupid question!

WOODMAN Michael
Jan 3, 2022, 2:59:58 AM1/3/22
to tvb-...@googlegroups.com
Hi
From the figure it is hard to say which is stable or unstable. Technically the eigenvalues of the Jacobian at a fixed point should have negative real parts for it to be stable, otherwise it is unstable. You can do it numerically in the case
of the WC model.
When you ask about FC appearing, I would guess you mean functional connectivity and specifically a fit between FC and SC as in some papers? If so this is not the bifurcation diagram to look at, rather you should do multiple simulations with varying
values of e.g. coupling strength and plot the variation of the SC-FC fit over the parameter value. This will give you an idea where the relevant bifurcation occurs.
Cheers,
Marmaduke
Looking forward for your reply! Please forgive my stupid question! <wilson-cowan.png>--
You received this message because you are subscribed to the Google Groups "TVB Users" group.
To unsubscribe from this group and stop receiving emails from it, send an email to tvb-users+...@googlegroups.com.
To view this discussion on the web visit https://groups.google.com/d/msgid/tvb-users/b054c17a-b130-4b77-9348-2d7dd3f1e2c5n%40googlegroups.com.
<wilson-cowan.png>
Wer White
Jan 3, 2022, 5:57:52 AM1/3/22
to TVB Users
Thanks,
Marmaduke.
'
From the figure it is hard to say which is stable or unstable. Technically the eigenvalues of the Jacobian at a fixed point should
have negative real parts for it to be stable, otherwise it is unstable.
You can do it numerically in the case
of the WC model.'
However, the arrows in the figure are from blue circle to yellow circle, does this can imply that the blue one is unstable and yellow one is stable?
'
When you ask about FC appearing, I would guess you mean functional
connectivity and specifically a fit between FC and SC as in some papers?
If so this is not the bifurcation diagram to look at, rather you
should do multiple simulations with varying
values of e.g. coupling strength and plot the variation of the SC-FC
fit over the parameter value. This will give you an idea where the
relevant bifurcation occurs.
'
Yes, i mean that
the emerging RSNs best account for the FC measured
experimentally using fMRI when the dynamical state of the net work is at the edge of a critical point , so does the crtical point is about the
bifurcation parameters but not the stable or unstable fixed point?
So how to decide the value and the concrete parameter of the critical point? Just to
do multiple simulations with varying
values and find the best one?

John Griffiths
Jan 3, 2022, 3:28:20 PM1/3/22
to TVB Users
Dear Wer,
Your questions are all very on-point on and not stupid in the slightest.
I think the concrete answer to your Q is yes: vary all combinations of global coupling and a few WC params and look for the combinations giving maximal match to empirical FC.
I will let Marmaduke confirm and elaborate on that as he sees fit.
Seems to me it might be useful to highlight a conceptual point here, which is why I'm jumping in:
In the comp neuro literature on whole-brain modelling of neuroimaging data using TVB and related tools, the idea of brain dynamics operating at a 'critical working point', 'close to a bifurcation', etc., is a major theme and broad consensus in the field (although specific instantiations of this in models of vary considerably). This is almost certainly an important insight about the basic principles of brain organization.
When you get into the details of working on this, as you are doing, you need to know that there are actually (at least) two versions of this idea; specifically the system which this idea is describing:
i) 'single node' / 'neural mass' / 'brain region' level
ii) 'whole brain' / 'whole network' level
In both of these cases, the ideas of bifurcations, critical working points, etc., all apply.
However, with a small number of exceptions*, bifurcations in the strict mathematically well-characterized sense are only known and/or well understood for the single-node case, such as the dynamical systems given by the Wilson-Cowan equations you have been studying in your phase plane analyses, the Fitzhugh-Nagumo equations (c.f. e.g. Figure 2a of Spiegler et al. 2016), the Reduced Wong-Wang model (c.f. Figs 18 and 19 in Sanz-Leon et al. 2015; phase flow diagram code here amongst other places), and of course the various Epileptor model variants.
The network-level sense of bifurcations in this context are usually associated with things like sharp changes in the simulated-empirical FC fit or the number of attractor states (as e.g. defined by a PCA, or by just listing of the number of distinct firing rate levels in the network, c.f. figs 2 and 3 in the paradigmatic study by Deco et al. 2013 ).
Importantly: i) and ii) are not mutually exclusive / independent of each other. That is, to achieve a good model of brain dynamics you will likely need to set node-level dynamics to a critical point (by tuning the neural mass model parameters), and also to set network-level dynamics to a critical point (by tuning network parameters like global coupling and possibly conduction delays). You will therefore often see in papers things something like a brute-force parameter sweeps over a combination of both local and global parameters, such as E-E and E-I coupling, Global coupling ('g'), and delays.
*Strictly speaking* once you are at the level of the whole-brain network model, the bifurcation structure of the individual neural mass models does not really apply, since they are now embedded in a larger complex system with many many more dimensions and degrees of freedom. *Practically speaking*, however, the node-level dynamics is almost always highly determinative of the overall system dynamics (in terms of e.g. presence of oscillations, oscillation frequency, firing rates, activity levels, etc. ); which is of course why we even bother to do things like look at single-node phase plane when our intention is to study whole-network behaviours.
So in summary - be on the look out for whether some described bifurcation behaviour you are reading about in a whole-brain modelling paper is talking about version i) or version ii), or some combination of them.
I hope this little exegesis doesn't muddy things for you too much. I do feel the distinction between node-level and network-level bifurcations and criticality isn't articulated as often as it probably should be given its importance to the modelling problems we are typically interested in.
Good luck with your work and have fun!
JG
*( namely something like the linear dynamical system dx/dt = Ax, where A is a macro-connectome and x is a vector of activity states )
Wer White
Jan 3, 2022, 7:46:16 PM1/3/22
to tvb-...@googlegroups.com
Thanks very much! Your letter really helps me! And I have another concrete question about 'node level
bifurcation', when i tune parameters e.g. x, and the hypothetical
threshold as x0, does it mean that when x<=x0, the fixed point(0,0),
i.e. resting state, E=I=0, is stable(unstable) and when x>x0, the
fixed point(0,0) is unstable(stable)?
John Griffiths <j.davidg...@gmail.com> 于2022年1月4日周二 04:28写道:
You received this message because you are subscribed to a topic in the Google Groups "TVB Users" group.
To unsubscribe from this topic, visit https://groups.google.com/d/topic/tvb-users/NIhEHtyLZ90/unsubscribe.
To unsubscribe from this group and all its topics, send an email to tvb-users+...@googlegroups.com.
To view this discussion on the web visit https://groups.google.com/d/msgid/tvb-users/a82aba2e-8eb1-493c-a581-c66e205fa7ben%40googlegroups.com.
Message has been deleted
Wer White
Jan 4, 2022, 10:52:38 AM1/4/22
to tvb-...@googlegroups.com
Sorry to bothor you! I have some new questions about the bifurcation parameter of neural mass model.
I find there may many parameters of the model can effect the phase plane of the neural mass. So how to decide which one to be the critical parameter?
And does it mean that there are many possibilities of the critical point and its value? So how to decide the best one? Just to test the fit of efc and sfc?
The most puzzled one is how to find the value, it's so hard to get it just from the phase plane.
* Above questions are all based on the 'node level'.
Looking forward to your reply! Thank you very much!
I find there may many parameters of the model can effect the phase plane of the neural mass. So how to decide which one to be the critical parameter?
And does it mean that there are many possibilities of the critical point and its value? So how to decide the best one? Just to test the fit of efc and sfc?
The most puzzled one is how to find the value, it's so hard to get it just from the phase plane.
* Above questions are all based on the 'node level'.
Looking forward to your reply! Thank you very much!
Wer White <wer.wh...@gmail.com> 于2022年1月4日周二 08:46写道:
Thanks very much! Your letter really helps me! And I have another concrete question about 'node level bifurcation', when i tune parameters e.g. x, and the hypothetical threshold as x0, does it mean that when x<=x0, the fixed point(0,0), i.e. resting state, E=I=0, is stable(unstable) and when x>x0, the fixed point(0,0) is unstable(stable)?
--
You received this message because you are subscribed to the Google Groups "TVB Users" group.
To unsubscribe from this group and stop receiving emails from it, send an email to tvb-users+...@googlegroups.com.
To view this discussion on the web visit https://groups.google.com/d/msgid/tvb-users/83c89ee1-d6f0-4a61-aa05-46dba9cc3b96n%40googlegroups.com.
Reply all
Reply to author
Forward
0 new messages